Modelo de ancestría múltiple desde la Teoría de Linajes
Modelo de ancestría múltiple desde la Teoría de Linajes
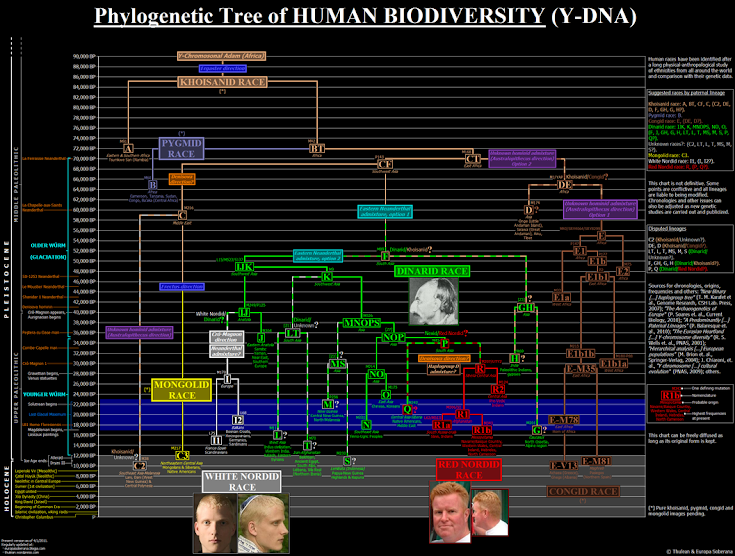
La comprensión de la diversidad humana y sus orígenes es un campo complejo que involucra múltiples disciplinas, entre ellas la genética molecular, la genética de poblaciones, la antropología física, la paleoantropología y la arqueología. En este contexto, la Teoría racial de linajes propone un modelo evolutivo basado en el análisis de haplogrupos paternos y maternos, junto con los rasgos morfológicos y antropológicos, para identificar subespecies o razas primigenias dentro de la especie humana. Esta teoría sostiene que dichas subespecies surgen por procesos naturales de mutación y selección, reflejando linajes genéticos claramente diferenciados y homogéneos en sus características físicas y biológicas.
Paralelamente, los modelos genéticos modernos, fundamentados en el estudio del ADN antiguo, han revelado que las poblaciones humanas actuales derivan de mezclas complejas y dinámicas de diversas poblaciones ancestrales. Estos modelos muestran un panorama multifactorial y en constante cambio, que puede parecer ambiguo o impreciso para algunas perspectivas paleoantropológicas clásicas, que buscan definiciones más claras y consistentes de las categorías raciales y subespecies.
Propongo un enfoque integrador, que busca armonizar las bases genéticas y filogenéticas del estudio de la evolución humana con los criterios morfológicos y de linajes ancestrales de la teoría racial. A través de un análisis cuidadoso, se explorarán las correspondencias aproximadas entre las poblaciones ancestrales identificadas genéticamente y las categorías tradicionales establecidas en la paleoantropología, con el fin de ofrecer una visión más completa y coherente de la evolución y diversidad humana
A diferencia de los modelos puramente poblacionales que priorizan la mezcla continua entre grupos, este enfoque parte de la premisa de que, en ciertos períodos de la prehistoria, existieron núcleos humanos relativamente aislados, sometidos a presiones ambientales únicas, que generaron tipos humanos diferenciados y estables en el tiempo. Cada uno de estos tipos estaría asociado a combinaciones específicas de haplogrupos maternos (mitocondriales) y paternos (cromosoma Y), así como a patrones consistentes de rasgos morfológicos y comportamentales.
Estos grupos originarios, no sólo pueden reconstruirse mediante el análisis genético de los haplogrupos modernos, sino también observando la coherencia morfológica de ciertos pueblos tradicionales aún existentes o documentados, así como restos fósiles bien conservados. El aislamiento reproductivo, la endogamia y la selección sexual habrían jugado un papel crucial en la fijación de estos rasgos, dando lugar a verdaderas subespecies humanas o razas primigenias.
Aunque muchas de estas poblaciones originales se hayan mezclado a lo largo del tiempo —ya sea por migraciones, guerras o procesos coloniales—, es posible detectar la huella de estos linajes fundacionales en el ADN y en la expresión física de los grupos humanos actuales.

Raza Congoide: Aparecen de la hibridación de khoisanidos con poblaciones arcaicas locales (posiblemente Homo Erectus u Homo Ergaster) y mutaciones adaptativas al fuerte clima subsahariano.
Raza Australoide: Hibridación entre Proto-khoisanido, Neandertal, Denisovano y Homínido Arcaico Desconocido. Viviendo en la comodidad y la baja existencia del ambiente.
Raza Armenida: Raza que surge de la diversificación de Proto-khoisanidos, con ausencia de herencia arcaica notoria. Moldeados por la región montanyosa de Medio Oriente durante la ultima glaciación (nariz grande y rostro aerodinámico), también por la agricultura (estructura facial más pobre).
Raza Nórdico Roja: Adaptación al clima extremadamente ártico de Asia Central que se dividió de la Raza Armenida hace aproximadamente 40.000 anyos, posible relación con la Raza Nórdico Roja; alta feomelanina por el frío extremo y rasgos robustos, cuerpos compactos.
Raza Mongoloide: Diversificación de un grupo africano primitivo (Proto-khoisanido) que llego a Asia; producto de adaptación al frio euroasiático (frio y seco) reforzando sus herencias arcaicas (Hombre de Denisova, Neandertal y posiblemente Hombre de la Caverna de los Ciervos).
Raza Cromagnoide: Resultado de migración progresiva (Armenidos del Levante → Norte de Mesopotamia → Cáucaso Sur → Cruce del Cáucaso → Cáucaso Norte → Borde norte del Mar Negro → Cuenca del Dniéster y del Danubio inferior → Europa Oriental → Europa Central → Europa Occidental → Cro-Magnon clásico), adaptación al frío durante glaciación de Würm (Ley de Allen y Bergmann), mixtura con neandertales (remanente de ~3%) y posterior gracilización con la Revolución Neolitica.
Se propone que la especie Homo sapiens se diversificó tempranamente en varios grupos o "linajes" fundamentales. Estos linajes habrían evolucionado en distintos contextos geográficos y ambientales, desarrollando características morfológicas propias.
A diferencia de la visión genética moderna —que ve la variación humana como un continuo sin límites raciales definidos— esta teoría postula que existen grupos bien diferenciados desde el Paleolítico, con persistencia parcial hasta hoy en ciertas regiones.
Cada uno de estos linajes se considera ancestral o representativo de ciertas poblaciones humanas, tanto históricas como contemporáneas, basándose en rasgos físicos, proporciones craneales y otras características visibles.
Estos linajes, presentados a continuación con su abreviatura correspondiente, son:
NR: Nórdico Rojo
ARM: Armenido
MONG: Mongoloide
KHO: Khoisánido
CONG: Congoide
PIG: Pigmeo
AUS: Australoide
Composiciones ancestrales generales con grupos intermedios
CRO (Cazadores Recolectores Occidentales): 100% CROG (posibles residuo MONG o CONG) CRE (Cazadores Recolectores del Este): ~75% AEN + ~25% CRO AEN (Ancestrales Euroasiáticos del Norte): 70% NR + 20% CROG + 10% MONG CRC (Cazadores-recolectores del Cáucaso): ~60% linaje cercano a AEN + ~40% ARM CRS (Cazadores Recolectores Escandinavos): ~55% CRO + ~35% CRE + ~10% AEN Botai (Kasajistán, 3700-3100 a.C., Domesticadores de Caballos): ~60% AEN + ~30% CRE + ~10% MONG Okunevo (Altái, Edad del Bronce temprano, 4.000): ~50% AEN + ~30% MONG + ~20% ARM Mesolíticos del Norte: ~60% CRO + ~40% CRE Irán Neolítico: ~50% ARM + ~50% CRC Natufios: ~50% ARM + ~40% CRAO + ~10% CRO CRA: (Cazadores Recolectores Anatolios): ~60% CRO + ~20% ARM + ~20% CRC Anatolia Neolítico: ~60% CRA + ~30% ARM + ~10% CRC Bactria-Margiana (Asia Central 2200-1700 a.C.): ~58% Irán Neolítico + ~29% Anatolia Neolítico + 11% Yamnaya ~2% Hoabinhiano Levante Neolítico: ~67% Natufios + ~33% Anatolia Neolítico PAE (Primeros Agricultores Europeos): ~85% Anatolia Neolítico + ~15% CRO Yamnaya: ~50% CRE + ~50% CRC CCC (Cultura de la Cerámica Cordada):~74% Yamnaya + ~23% Anatolia Neolítico + ~3% WHG AIN (Ancestros Indios del Norte) ~70% Yamnaya + ~30% Irán Neolítico AIS (Ancestros Indios del Sur): ~55% Irán Neolítico + ~45% AUST SA (Sudeste Asiático): ~90% MONG + ~10% AUS NA (Negrítido Austronesio): ~40% AUS + ~60% MONG PA (Papúes ancestrales): ~100% AUS MA (Melanesios ancestrales): ~80% AUS + ~20% MONG MO (Mixtura Oceánica): ~70% AUS + 30% MONG Hoabinhianos (Vietnam, 4-8000 a.C.): ~75% AUS + ~25% MONG
Desglose detallado en linajes ancestrales puros y porcentajes precisos
CRO: 100.0% CROG
AEN: 70.0% NR + 20.0% CROG + 10.0% MONG
CRC: 42.0% NR + 40.0% ARM + 12.0% CROG + 6.0% MONG
CRS: es Botai: 57.75% NR + 24.00% CROG + 18.25% MONG
Okunevo: 35.00% NR + 35.00% MONG + 20.00% ARM + 10.00% CROG
Natufios: 62.00% ARM + 28.00% CONG + 10.00% CROG
CRA: 62.40% CROG + 28.00% ARM + 8.40% NR + 1.20% MONG
Anatolia Neolítico: 50.80% ARM + 38.64% CROG + 9.24% NR + 1.32% MONG
Bactria-Margiana: 57.50% ARM + 20.10% NR + 17.60% CROG + 4.80% MONG
Levante Neolítico: 58.30% ARM + 19.45% CROG + 18.76% CONG + 3.05% NR + 0.44% MONG
Yamnaya: 47.25% NR + 26.00% CROG + 20.0% ARM + 6.75% MONG
NA: 60.00% MONG + 40.00% AUS
MA: 80.00% AUS + 20.0% MONG
MO: 70.00% AUS + 30.0% MONG
ASO: 45.50% NR + 41.5% MONG + 13.0% CROG
BA: 52.00% MONG + 20.0% ARM + 14.0% CROG + 14.0% NR
PS: 78.75% MONG + 16.28% NR + 4.65% CROG + 0.33% ARM
Eskimo-Aleut: 74.74% MONG + 15.93% NR + 6.05% CROG + 3.28% ARM
Paleoárticos: 71.0% MONG + 22.05% NR + 6.3% CROG + 0.65% ARM
AN: 51.5% MONG + 20.0% ARM + 13.0% CROG + 10.5% NR
CRAC: 100.00% PIG
NIA: 100.00% CONG
AO: 100.00% CONG
CRAO: 70.00% CONG + 30.00% ARM
Iberomaurisianos: 55.00% ARM + 25.00% CONG + 20.00% CROG
Hadza: 60.00% CONG + 25.00% KHO + 15.00% ARM
Protobereber: 52.90% ARM + 29.32% CROG + 12.50% CONG + 4.62% NR + 0.66% MONG
Preguntas Frecuentes: Aclaraciones sobre Linajes Humanos y Composición Genética
Dada la complejidad del modelo de linajes y la diversidad de grupos humanos implicados, es natural que surjan interrogantes respecto a las poblaciones, sus mezclas, sus rasgos y su clasificación dentro del marco propuesto. A continuación, se abordarán cuestiones relevantes que pueden emerger tanto desde temáticas periféricas como desde aspectos internos al sistema aquí expuesto.
En esta sección se abordan de forma clara y directa las preguntas más recurrentes en torno a la exégesis de los linajes, la interpretación de sus mezclas, y la identificación de grupos humanos pasados y presentes dentro de este marco teórico.
¿Cómo llego la Influencia Armenida en Poblaciones Norteméricanas?
Esto se sabe gracias al rastreo haplogrúpico y a los estudios genómicos de poblaciones. Aunque algunas teorías han llegado a conclusiones muy extremas —como utilizar estos datos para apoyar la hipótesis solutrense—, deben ser tomadas con suma cautela. El haplogrupo X mitocondrial habría surgido hace unos 30.000 años, a partir de una rama del haplogrupo N, en la región del Cáucaso.
Se ha documentado que migraciones procedentes de Medio Oriente alcanzaron Siberia y Asia Central hace aproximadamente 30.000 años, influyendo en poblaciones como los ANE (Ancient North Eurasians). Sin embargo, estas migraciones se intensificaron durante el Pleistoceno final, hace unos 12.000 años, coincidiendo con la gracilización asociada al cambio climático.
El haplogrupo X2 se habría diversificado en Siberia, dando origen al subclado X2a, presente en algunos grupos beringios ancestrales.
Asimismo, considerando que el aporte ANE en los pueblos nativos de Norteamérica alcanza solo un ~15–25%, en comparación con otras poblaciones amerindias que presentan un ~35–50%, se puede inferir que existió una posible influencia de poblaciones siberianas con un aporte amerindio considerable.

¿La Revolución Neolitica provoco la gracilización de los Cromagnoides?
Sí. El proceso de gracilización observado en las poblaciones europeas está estrechamente vinculado a la transición hacia la agricultura iniciada en el Próximo Oriente entre el 9.000-7.000 a.C., con el uso del trigo, cebada y diversas legumbres. Desde estas regiones. las innovaciones agrícolas se expandieron hacia Anatolia, donde los grupos locales de cazadores-recolectores adoptaron progresivamente estas prácticas, generando una población de anatolios neolíticos. Mantenían una fuerte impronta cromagnida, por lo que no pueden considerarse completamente armenoides.
Desde Anatolia, la agricultura de difudió hacia Europa por dos grandes rutas: la mediterránea y la danubiana. Los agrilcutores coexistieron con las poblaciones mesolíticas, generandose un mestizaje biológico y cultural: mientras los cazadores-recolectores incorporaban gradualmente técnicas agrícolas, los agricultores también adaptaban saberes locales de subsistencia.
Con el tiempo, entre el 5500-4000 a.C. surgieron asentamientos agrícolas estables, en lugares como los Balcanes o Europa Central. Posteriormente, hacia el 4.000-3.000 a.C., el modelo agrícola terminó de consolidarse en las regiones septentrionales del continente.
Este cambio en el modo de vida supuso una profunda transformación en la morfología cromagnida. La dieta agricola, abundante en calorías gracias a cereales como trigo, cebada y mijo, pero comparativamente mucho más pobre en proteínas y micronutrientes que las dietas de los cazadores-recolectores, condujo a una diminución en la talla promedio (10 cm menos o mas), menor densidad ósea y reducción de robustez de los rasgos craneofaciales (cráneo menos grande, arcos superciliares menos marcados, mandíbula y pómulos más estrechos, etc.), la vida sedentaria implicó un descenso en las demandas físicas cotidianas.
En conjunto, estos factores explican la transición desde poblaciones tipo cromagnidas a los grupos Nórdico Blanco Europeos posteriores.

Comentarios
Publicar un comentario